Значение слова «энтодерма»

У первичноротых энтодерма выстилает полость первичной кишки, из неё образуется средняя кишка и все её добавочные железы.
У вторичноротых образует внутренний слой кишечной трубки.
У позвоночных из энтодермы развивается слизистая оболочка всего кишечника и связанные с ним железы (печень, поджелудочная железа и др.).
У рыб, кроме того — плавательный пузырь и внутренние жабры, а у высших позвоночных — лёгкие.
энтоде́рма
1. биол. внутренний зародышевый листок у многоклеточных животных, из которого образуются кишечный тракт и связанные с ним железы ◆ Личинка принимает скоро характер настоящей гаструлы и состоит уже из эктодермы — наружного слоя и энтодермы — внутреннего; она имеет форму мешка с отверстием на нижнем конце. И. И. Мечников, «Лекции о сравнительной патологии воспаления», 1892 г. (цитата из НКРЯ)
2. биол. внутренний клеточный слой стенки тела кишечнополостных животных
Делаем Карту слов лучше вместе
 Привет! Меня зовут Лампобот, я компьютерная программа, которая помогает делать Карту слов. Я отлично умею считать, но пока плохо понимаю, как устроен ваш мир. Помоги мне разобраться!
Привет! Меня зовут Лампобот, я компьютерная программа, которая помогает делать Карту слов. Я отлично умею считать, но пока плохо понимаю, как устроен ваш мир. Помоги мне разобраться!
Спасибо! Я обязательно научусь отличать широко распространённые слова от узкоспециальных.
Насколько понятно значение слова треножник (существительное):
Энтодерма возникла в ходе эволюции раньше эктодермы
Автор
Редактор
Исследование активностей всех генов в ходе развития зародыша круглого червя Caenorhabditis elegans выявило новые онтогенетические детали. Оказывается, внутренний зародышевый листок — энтодерма — приобретает свои специфические особенности раньше внешнего листка — эктодермы. Такой же порядок развития зародышевых листков обнаружился и у других видов животных. Кроме того, гены, активные в энтодерме были в среднем «старше», чем гены, работающие в других зародышевых листках. Это указывает на более древнее происхождение энтодермы в ходе эволюции по сравнению с эктодермой.
В эмбриональном развитии даже довольно непохожих организмов можно выделить сходные стадии. Оплодотворенная яйцеклетка делится, образуя комок клеток, в котором формируется полость (комок клеток с полостью называется бластулой). Часть поверхности полого шарика начинает впячиваться внутрь (этот процесс называется гаструляцией), и образуется нечто вроде двухслойного мешочка (который называется гаструлой). Его внешний слой становится зародышевым листком эктодермой, из которого образуются покровы тела и чувствительные клетки. Из внутреннего слоя, который называется энтодермой, формируется пищеварительная система и другие внутренние органы. Животные, развивающиеся только лишь их этих двух зародышевых листков — энтодермы и эктодермы, называются двухслойными. К ним относятся кишечнополостные и гребневики — достаточно просто устроенные организмы с радиальной симметрией, а также губки — еще проще устроенные животные вообще без симметрии и настоящих тканей.
У более сложно организованных животных в ходе эмбрионального развития возникает и третий зародышевый листок — мезодерма. Это выпячивание энтодермы, образующее самостоятельный слой. Из мезодермы берут начало мышечные клетки и скелет. Животные, которые развиваются из трех зародышевых листков — двусторонне-симметричные: у них есть правая и левая стороны тела, в отличие от, например, медузы. Насчет того, что мезодерма появилась в эволюции позже энтодермы и эктодермы, сомневаться не приходится — всё же двухслойные животные устроены проще, чем трехслойные, и возникли раньше их. А в статье, недавно опубликованной в журнале Nature [1], приводятся данные, из которых следует, можно сказать, революционный вывод: энтодерма, оказывается, возникла раньше эктодермы. Казалось бы, эти два слоя — две стороны одной гаструлы (рис. 1). Могло ли быть так, что один из них возник в ходе эволюции раньше?
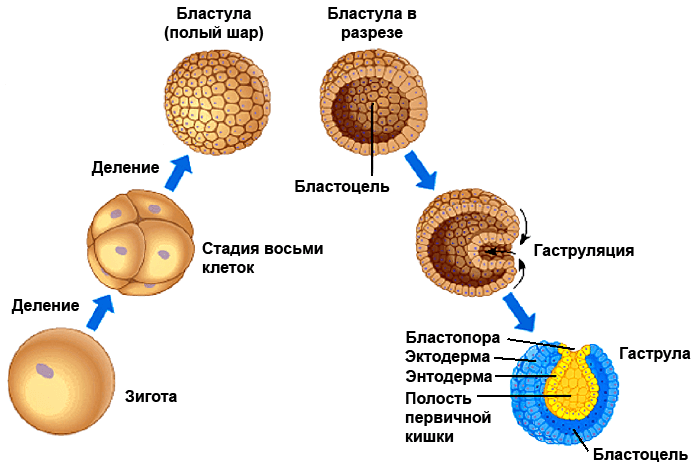
Рисунок 1. Стадии развития зародыша от оплодотворенной яйцеклетки до гаструлы. Рисунок с сайта studopedia.org (адаптирован).
Ученые решили изучить процессы формирования зародышевых листков, ориентируясь на данные по активности генов. Чтобы не мелочиться и не упустить чего-то важного, исследователи оценили активности всех генов, которые работают на протяжении всего эмбрионального развития круглого червя (нематоды) от стадии пяти клеток до взрослой личинки. Этот червь — Caenorhabditis elegans — один из самых подробно изученных организмов. Например, точно известна судьба каждой из пяти первых клеток зародыша. Это стало возможным в том числе и благодаря удивительно аккуратному развитию нематоды: любая взрослая гермафродитная особь состоит ровно из 959 клеток (для этого червя характерна эутелия). К тому же небольшое число клеток позволяет в подробностях изучать строение целого организма. К примеру, нервная система червя, состоящая из 302 нейронов, описана с точностью до каждой связи.
Используя относительно новый метод исследования РНК индивидуальных клеток (CEL-seq)*, ученые получили профили экспрессии всех генов червя в зависимости от времени и расположения в развивающемся зародыше. Методом главных компонент удалось выделить группы генов, специфично экспрессирующихся в эктодерме, энтодерме и мезодерме. Гены, активные в мезодерме, ожидаемым образом начинали транскрибироваться позже других групп. Неожиданным же оказалось то, что экспрессионная программа энтодермы запускалась раньше, чем активировались гены эктодермы. Получается, что эти зародышевые листки приобретают свои характерные черты не одновременно.
* — Подходы к секвенированию генома и транскриптома индивидуальной клетки описаны в работе «Секвенирование единичных клеток (версия — Metazoa)» [2], а транскриптомов тысяч индивидуальных клеток сразу — в статье «Разработан метод анализа экспрессии генов на уровне индивидуальных клеток» [3]. — Ред.
Чтобы проверить, не является ли такой порядок развития зародышевых листков особенностью одних лишь нематод, ученые исследовали профили экспрессии генов и у других организмов: губки (Amphimedon queenslandica), актинии (Nematostella vectensis) и лягушки (Xenopus tropicalis). У всех этих животных тенденция была такой же: ортологи генов, экспрессируемых у нематоды в энтодерме, «включались» в ходе эмбрионального развития первыми (рис. 2). Получается, что у зародышей разных животных энтодерма появляется раньше эктодермы. Но означает ли это, что и в эволюции энтодерма возникла раньше?
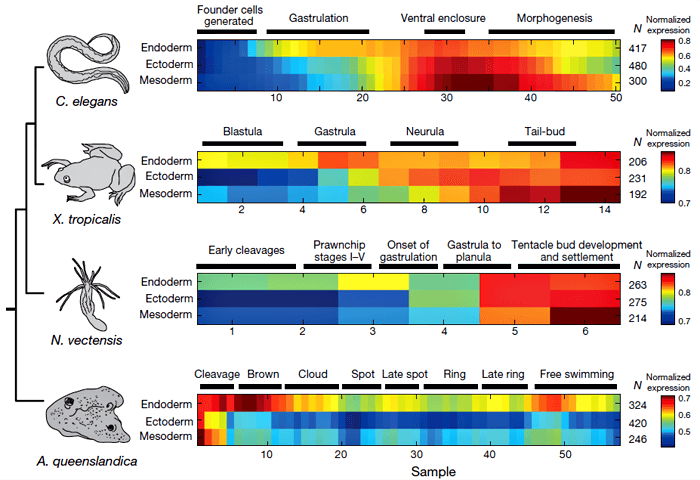
Рисунок 2. Гены, активные в энтодерме, «запускаются» у разных видов животных раньше, чем гены эктодермы и мезодермы. В последнем ряду данных, относящемся к губке, показано, что гены эктодермы начинают экспрессироваться не позже энтодермальных. Это связано с тем, что эктодермальные гены у губок проявляют активность только посредством материнских транскриптов, сохраняющихся в яйцеклетке. Рисунок из [1].
Раз уж ученые определили гены, активные в разных зародышевых листках, то биоинформатическими методами стало возможным определить и эволюционный возраст этих генов. В общих чертах это делается следующим образом: берется филогенетическое древо с достаточно разнообразными организмами и для каждого исследуемого гена определяется, у кого и когда впервые возникла последовательность, в достаточной степени похожая на этот ген. Так ученые выяснили, что гены, экспрессирующиеся в энтодерме червя Caenorhabditis elegans, в среднем старше генов, активных в других зародышевых листках. Это веский довод в пользу того, что энтодерма возникла в ходе эволюции раньше эктодермы.
Такое открытие чрезвычайно важно для теорий возникновения многоклеточности. Согласно одной из них, однажды клетки со жгутиками, напоминающие современных хоанофлагеллят, «решили» объединиться в шарообразную структуру с пищеварительной полостью внутри. Исследователи сравнили гены представителя этой группы одноклеточных (Monosiga brevicolli) с генами, избирательно экспрессирующимися в трех зародышевых листках нематоды. Оказалось, что доля ортологов генов Monosiga выше среди генов, активных именно в энтодерме. Это неудивительно, учитывая, что гены, работающие в энтодерме, отвечают в основном за функции, связанные с питанием: производство энергии, метаболизм и транспорт, — то есть вопросы первоочередной важности для любого организма.
Так что, принимая во внимание порядок возникновения слоев, можно представить такую модель формирования многоклеточных:
Все три слоя дали организму невероятные возможности, и трехслойный дизайн позволил возникнуть большому разнообразию сложных организмов. Но началось всё с энтодермы — самого важного для жизни зародышевого листка.
Что входит в энтодерму
Отечественные вузовские курсы ботаники, программы которых весьма близки, обязательно включают материал о закономерностях внутреннего строения вегетативных органов растений, о разнообразии анатомии органов у растений различных таксонов. При обучении ряду специальностей (биология, агрономия, фармация и др.) одной из задач курса ботаники является выработка у студентов навыков самостоятельного анализа срезов вегетативных органов на лабораторных занятиях.
Излагая теоретический материал по анатомии вегетативных органов растений, преподаватель, в частности, должен давать представления об эндодерме и экзодерме. Трудность заключается в том, что названные понятия трактуются в учебной ботанической литературе неоднозначно.
Нами установлено, что в трактовке экзодермы и эндодермы просматриваются две тенденции.
Часть авторов склонны рассматривать эндодерму и экзодерму как особые ткани растений, слагающие слои первичной коры [2, 4].
Другая точка зрения заключается в том, что экзодерма и эндодерма считаются анатомо-топографическими зонами органа.
И.И. Андреева и Л.С. Родман [1], рассматривая классификацию растительных тканей и характеризуя их группы, экзодерму и эндодерму в числе тканей не называют. Описывая строение корня, они разделяют первичную кору на экзодерму, мезодерму и эндодерму, никак их не квалифицируя. Упоминается также и экзодерма стебля, по выражению авторов, представленная колленхимой. Таким образом, экзодерма и эндодерма явно трактуются как топографические зоны органов.
Зачастую авторы, придерживающиеся указанной (второй) точки зрения, при описании анатомических препаратов не называют ткани, составляющие топографические зоны экзодерму, мезодерму и эндодерму, а описывают клетки (например: клетки мезодермы крупные, тонкостенные и пр.).
Признаки экзодермы и эндодермы, свойственные им как растительным тканям, приводятся ниже. В общих чертах они характеризуются в учебниках более или менее одинаково, вне зависимости от того, рассматриваются ли экзодерма и эндодерма как ткани либо как топографические зоны.
Экзодерма и эндодерма имеют плотно сомкнутые клетки со специфическими утолщениями клеточных стенок. У эндодермы первоначально формируются утолщения в виде поясков Каспари. Впоследствии у ряда растений утолщения стенок клеток эндодермы приобретают иную конфигурацию. Пояски Каспари не пронизываются плазмодесмами, лигнифицируются, инкрустируются липофильными веществами и перекрывают радиальный апопластный транспорт через эндодерму. Перемещение веществ в таких условиях возможно лишь по симпласту, поэтому становится регулируемым. П. Зитте с соавтора-
ми [2] указывают, что клетки экзодермы также имеют пояски Каспари. Вследствие описанных особенностей строения, а также топографии, экзодерме и эндодерме присущи определённые функции, которые наиболее ярко проявляются у этих тканей в зоне поглощения корня. Экзодерма корня, располагающаяся на периферии первичной коры, пропуская растворы минеральных солей из эпиблемы в первичную кору, препятствует их обратному току, а эндодерма корня, выстилающая первичную кору изнутри, пропуская минеральные соли в стелу, препятствует выходу их из центрального цилиндра. Таким образом, развитие экзодермы и эндодермы с описанными выше особенностями строения клеток свидетельствует о наличии поглощения органом веществ извне и их регулируемого радиального транспорта в толще органа.
Мы считаем необходимым пояснять студентам, что характеристики тканей часто не являются статичными и зависят от того, в теле какого растения, в каком органе и в какой части органа они располагаются, а также от степени развитости («степени зрелости») органа. Так, эндодерма корня может проходить три стадии развития:
— формирование поясков Каспари;
— суберинизация клеточных стенок;
— значительное утолщение стенок большинства клеток (кроме пропускных клеток), сопровождающееся гибелью их протопластов.
В проанализированных нами отечественных учебниках ботаники пояски Каспари у клеток экзодермы не описываются, барьерная функция экзодермы не обсуждается, а констатируется лишь её роль в качестве механического покрова корня. Как указывалось выше, Л.И. Лотова [4] даже относит экзодерму корня к покровным тканям. П. Зитте с соавторами [2] в некоторых случаях именуют периферическую ткань кортекса корня гиподермой.
Эндодерму и экзодерму в учебниках следовало бы анализировать на серии поперечных срезов корня в зоне всасывания и в зоне проведения, что позволило бы дать представления обо всех стадиях их развития. Однако в рассматриваемых отечественных учебниках описывается лишь строение корня ириса, эндодерма и экзодерма которого находятся на завершающих стадиях своего развития.
Внутренний слой первичной коры стебля часто описывается как «крахмалоносное влагалище», поскольку в его клетках обнаруживаются многочисленные крахмальные зёрна. Пояски Каспари и прочие характерные утолщения клеточных стенок здесь не обнаруживаются. Крахмалоносному влагалищу приписывается функция обеспечения определённого положения стебля в пространстве, т.к. крахмальные зёрна в его клетках играют роль статолитов. В отечественных учебниках названия «крахмалоносное влагалище» и «эндодерма» обычно приводятся как синонимы. В контексте нашей трактовки экзодермы и эндодермы как тканей, имеющих вполне определённые функции, и введения понятий «экзокортекс» «мезокортекс» и «эндокортекс» крахмалоносное влагалище следует рассматривать как особую, отличную от эндодермы, ткань, составляющую эндокортекс стебля. Что касается эндодермы, то в некоторых стеблях (например, в стеблях корневищ) она может развиваться, что свидетельствует о наличии в таких стеблях радиального симпластного транспорта веществ между кортексом и стелой. Экзодерма для стебля не характерна.
При трактовке эндодермы и экзодермы как растительных тканей введение топографического понятия «мезодерма» лишено смысла и его нужно избегать.
При анализе и описании среза органа удобно сначала формально различать подзоны кортекса. Далее следует обозначать ткани, их слагающие. В заключение необходимо устанавливать и описывать особенности каждой ткани, её клетки.
Выводы
1. Для преодоления имеющихся в учебниках разночтений и удобства анализа срезов вегетативных органов растений в понятийный аппарат курсов ботаники полезно ввести топографические понятия «экзокортекс», «мезокортекс» и «эндокортекс», обозначающие подзоны кортекса (первичной коры).
2. Целесообразно квалифицировать экзодерму и эндодерму не как анатомо-топографические зоны органов, а как ткани, проходящие в процессе своего развития и функционирования различные стадии, характеризующиеся определёнными структурными и функциональными особенностями.
Список литературы
Рецензенты:
Бессчетнов В.П., д.б.н., профессор, зав. кафедрой лесных культур ФГОУ ВПО «Нижегородская государственная сельскохозяйственная академия» Министерства сельского хозяйства РФ, г. Нижний Новгород.


